MORE NEANDERTAL INSIGHTS:
 Table 1 taken
from:
Table 1 taken
from:
KRINGS, DNA sequence
of the mitochondrial
hypervariable region II from
the Neandertal type specimen,
Proc. Natl.
Acad. Sci. USA,
Vol. 96, pp. 5581–5585, May
1999.
This article can be found and accessed for free at
(in html or in PDF):
Figure 1. Illustration of Neandertal Man ,
John Gurche/National Geographic.
A fragment of the original article that Lindahl reviews in the next space (the first mtDNA comparison of a neandertal bone) can be found for free at:
http://mendel.uab.es/biocomputacio/treballs98-99/Villatoro/artcell.htm
Tomas Lindahl, Facts and
Artifacts of Ancient DNA, Cell, Vol. 90, 1–3, July 11, 1997
Excerpts:
“Neandertals, named after
the German valley where these fossils were first discovered, were about 30% larger
than an average modern man and of great muscular strength. They had low
foreheads, protruding brows, and large noses with broad nostrils and were meat
eaters.
The paucity of
the fossil record has not allowed a direct resolution of this important problem,
although recent morphological studies of the nasal cavity of Neandertals favor
the alternative that they represented a distinct and separate species, Homo
neanderthalensis (Schwartz and Tattersall, 1996).
The small
amount of sequence divergence observed in mitochondrial DNA (mtDNA) from
different contemporary human populations, especially in Europe, also indicates
a relatively recent origin of Homo sapiens without significant admixture
of ancient Neandertal sequences… (Torroni, A., Lott, M.T., Cabell, M.F., Chen,
Y.-S., Lavergne, L., and Wallace, D.C. (1994). Am. J. Hum. Genet. 55,
760–776.)”
“Neandertals,
however, seem to have been more similar to modern humans than to apes in having
a low species-wide mtDNA diversity. In the case of humans, the low genetic
diversity seen in both mtDNA and nuclear DNA sequences is likely to be the
result of a rapid population expansion from a population of small size, often assumed
to have been made possible by some cultural
or genetic innovation, such as use of a complex language”.
Fragment taken from:
Krings, A view of Neandertal genetic diversity, Nature
Genetics volume 26 •
october 2000, 144-146
“For many
years, the Neanderthals have been recognized as a distinctive extinct hominid
group that occupied Europe and western Asia. Our ongoing studies indicate that
the Neanderthals differ from modern humans in their skeletal anatomy in more
ways than have been recognized up to now. The purpose of this contribution is
to describe specializations of the Neanderthal internal nasal region that make
them unique not only among hominids but possibly among terrestrial mammals in
general as well. These features lend additional weight to the suggestion that
Neanderthals are specifically distinct from Homo sapiens”.
Abstract:
Schwartz, J. H. and I. Tattersall (1996). “Significance of some
previously unrecognized apomorphies in the nasal region of Homo
neanderthalensis.” Proc Natl Acad Sci U S A 93(20): 10852-4.
The complete
article can be accessed for free at (in PDF):
Neandertal and
other possible Nephilim “seeds” (from the artistical reconstructions):
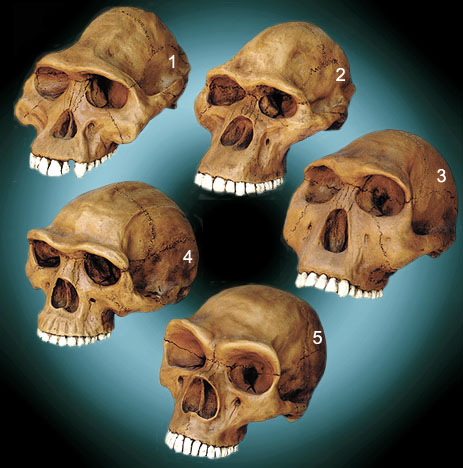
- Australopithecus
afarensis Cranium, 2. Australopithecus africanus Cranium,
- Homo habilis Cranium, 4. Homo erectus Cranium, 5. Neandertal
Cranium
Fragments of the history of this skulls:
Hominid
Series of Five Skulls
“It is from casts, photographs, published diagrams and text
describing these reconstruction's that sculptor Larry Williams has developed
this Hominid Series.
Isaac de la Payrere, from France, discovered stone tools
used by primitive men who, he claimed, lived in the time before Adam. In 1655
his findings and theory were greatly disapproved of and his books publicly
burned by Church authorities.
In 1856, three years before Darwin propounded his theory of
evolution, Neanderthal Man was discovered in Germany's Neander Valley near
Dusseldorf. The skeleton possessed a number of peculiar traits that defined
them as very ancient, including a low, narrow, sloping forehead, heavy eyebrow
ridges, and a deep depression at the root of the nose.
Between 1890 and 1892, Eugene Dubois (member of the Dutch
colonial army) found pieces of "Java Man," (later named Homo
erectus) that everyone agreed was more apelike than the Neanderthal. The
importance of H. erectus was vastly increased from 1927 to 1937 as more
than 40 similar fossils were found in limestone caves at Zhoukoudian, outside
of Beijing. Also found were thousands of stone tools and evidence that H.
erectus used fire. "Beijing Man" was somewhat like the Java erectus.
Homo erectus and Neanderthals were more manlike than apelike (Later
termed Australopithecus). Then, in South Africa, in 1924, Raymond Dart
discovered a apelike. It was followed by the discovery of similar apelike
creatures in Africa, with a brain only slightly bigger than a chimpanzee's. The
nose was flat. The jaw dominated the face and the mouth thrusted forward. But
the teeth were human like and it had a bit of a forehead. Most importantly, it
walked upright! Its spinal cored entered the brain not at the back of the head,
like a gorilla's, but at the bottom of the skull, suggesting bipedalism.
Although that didn't make it human, it allowed it to fall into the broader
category of "hominid."
In all of the fossil finds, however, there was no evidence of
where and when anatomically modern humankind first arose”.
Taken from: http://www.sculpturegallery.com/sculpture/hominid_series_of_five_skulls.html
Compare the skulls presented above
with the next (male and female skulls in artistical reconstructions also):

Human Male and Female Skulls
Taken from:
http://www.sculpturegallery.com/sculpture/human_male_skull.html
http://www.sculpturegallery.com/sculpture/human_female_skull.html
Does Homo neanderthalensis play a role in modern human ancestry? The mandibular evidence. Rak Y, Ginzburg A, Geffen E. Am. J. Phys. Anthropol., 2002 Nov;119(3):199-204:
“The specialized Neanderthal mandibular ramus morphology emerges as yet another element constituting the derived complex of morphologies of the mandible and face that are unique to Neanderthals. These morphologies provide further support for the contention that Neanderthals do not play a role in modern human biological ancestry, either through "regional continuity" or through any other form of anagenetic progression.”
Other fragment showing differences between
humans and neandertals:
“Whatever social contact may have occurred, Hublin and Fred Spoor of University College London conclude that Neandertals and early modern humans did not interbreed. Computerized- tomography scans of nine Neandertal temporal bones, which surround the inner ear, revealed small semicircular canals and a distinctive inner ear shape compared to modern humans. "The differences are comparable to those separating ape species," they said”... “The bone came from a 13- by 16-foot structure made of stalactite and stalagmite fragments. Built by Neandertals, its purpose is unknown.”
MARK BERKOWITZ
http://www.archaeology.org/9609/newsbriefs/neandertals.html
Archaeology Newsbriefs, Vol. 49 # 5
Sept/Oct 1996, Neandertal News
Excerpts of an excellent comment of the article of Schwartz and
Tattersall (1996) previously referred:
Laitman, J.
T., J. S. Reidenberg, Samuel Marquez, and Patrick J. Gannon. (1996). “What the nose knows: new
understandings of Neanderthal upper respiratory tract specializations.” Proc Natl
Acad Sci USA 93(20):
10543-5.
“Who were the
Neanderthals? This brief question has been at the heart of paleoanthropology
since the unearthing of the first Neanderthals almost a century and a half ago.
First found at sites in western Europe, such as the Neander ‘‘thal’’ (now
simply called “tal”) or valley in Germany, Gibraltar, Spy in Belgium, or La
Chapelle-aux-Saints in France, these peculiar remains continue to be an enigma
(1, 2). Were they mere variants of living human populations, somewhat different
so as to indicate a distinct ‘‘race,’’ or were they imbued with
sufficient uniquely derived characters (autapomorphies) so as to warrant their
own species, Homo neanderthalensis?... it is generally agreed that Neanderthals... inhabited a range including much of Europe, western Asia, and the Levant... what happened to them? Although
questions abound, definitive answers are few.
A
confounding problem in the search to understand the Neanderthals, as with
hominid(s)... in general, is that important aspects of their functional
morphology and underlying behaviors have gone largely underinvestigated. This,
in part, is because paleoanthropologists have had to focus upon the scant bony
remains available to them, remains which often do not reflect some major
biological systems. For example, although aspects of the dentition or
postcranial skeleton have been reconstructed meticulously, others, such as
features of the respiratory, digestive, or nervous systems have, more often
than not,
either been ignored or relegated to a relatively minor role in discussions.
The recent
observations by Schwartz and Tattersall (3), in this issue of the Proceedings,
on Neanderthal nasal apomorphies shed new light on the relationship of
Neanderthals to other hominids and add important new data for our understanding
of the functional morphology of a major physiologic system, the upper
respiratory tract. The authors’ primary interest is, of course, how these newly
described features will affect the interpretation of Neanderthal phylogeny. To
be fair, the position of Schwartz and Tattersall on this issue is well known
among anthropologists, with both recognized as ‘‘splitters’’ who see
Neanderthals as comprising a distinct species from either living or archaic Homo
sapiens (1, 4, 5). Their prior views notwithstanding, the evidence they
present is of value in its own right and will henceforth have to be factored
into any discussion of Neanderthal relationships.
The findings by
Schwartz and Tattersall regard specializations of internal nasal morphology.
This aspect of the nose has gone largely unexplored in our fossil ancestors.
This is because fragile internal nasal structures are often not reserved in
fossil hominids and because many aspects of the functional morphology
underlying the mammalian nose and associated paranasal sinuses remain poorly
understood. Although accounts of internal nasal morphology have been cursory, there
have been studies on external nasal dimensions of both extant and fossil
hominids (6–9). Indeed, the large size of Neanderthal external nasal
dimensions, and the significant differences they exhibit from those of extant
humans, has been commented upon and documented for some time (10–12).
Schwartz and
Tattersall now bring to the discussion three internal nasal features which have
been previously unrecognized and which they feel are autapomorphic to
Neanderthals: (i) an ‘‘internal nasal margin,’’ a medially projecting
rim of bone just within the anterior edge of the anterior nasal (piriform)
aperture; (ii)a pronounced medial swelling of the lateral nasal wall;
and (iii) a lack of an ossified roof over the lacrimal groove.
Should the
features observed by Schwartz and Tattersall indeed prove to be true
apomorphies, they would provide strong evidence to support the contention that
Neanderthals are different from extant or fossil Homo sapiens in seminal
ways. But in what ways? And what do distinctions in their nasal region reflect?
While focusing upon questions of Neanderthal systematics, Schwartz and
Tattersall have led us to ponder pivotal questions about how the Neanderthals
may have differed from us in aspects of their respiratory tract and constituent
behaviors.
The acquisition
and processing of oxygen and its byproducts is the primary mission of any
air-breathing vertebrate. Chewing, walking, reproducing, thinking are all fine,
but first one has to breathe. Anthropologists sometimes seem to forget this... Although...
some workers have attempted to
reconstruct the... aspects of our more elusive respiratory behaviors.
This work has focused largely on reconstructing the upper respiratory, or
aerodigestive, tract and has been principally concerned with establishing what
the overall positioning of structures such as the larynx, tongue, and pharynx
may have been like in hominid(s)... Our own studies (13–16) among others
(17–19) have shown that aspects of the external contour of the skull base are
intimately related to the topographic arrangement of aerodigestive tract
structures and can, in turn, serve as a guide to help reconstruct the anatomy
of the region... In essence, we have learned that the
basicranium is the ‘‘roof’’ of the upper respiratory tract and that it can
serve as a blueprint from... the ‘‘house’’ below... By
coupling these data on how upper respiratory tract structures can be
reconstructed with our growing knowledge of the comparative and functional
anatomy of this region in extant mammals (20, 21), we have begun to gain
insight into what the upper respiratory tract may have been like in…
australopithecines (22).
The
configuration of this region in Neanderthals has been the subject of thought
beginning, at least, with the great anatomist Sir Arthur Keith (23). The focus,
however, has usually not been on respiratory requirements and related behaviors
but, rather, on the ‘‘vocal tract’’ component. Although few studies wrestle
with the key issues of upper respiratory change, it seems as if almost everyone
has a word to say on what a Neanderthal’s vocal tract may have been like.
Although the
question of Neanderthal speech capabilities is indeed intriguing, this allure
has seemingly caused many to lose sight of what the area’s main functions were:
respiration and, secondarily, ingestion of food. The region did not evolve for
the sole purpose of vocalization, a fact often overlooked by many.
The
general picture of the Neanderthal upper respiratory tract that has emerged
over the last few decades by those attempting to reconstruct it has been of a
region which differed somewhat from that in living humans. Both our own (15,
16, 24) and other studies (17, 18, 25) have emphasized that some Neanderthals
(such as the ‘‘Classic’’ western European specimens) would have exhibited a
larynx slightly higher in the neck than that of modern humans, with these
Neanderthals having a more limited oropharyngeal segment with a greater portion
of the tongue occupying the oral cavity. When one factors in their large
external nose and sizable paranasal sinuses, the overall Neanderthal anatomy
suggests a group that relied more heavily upon the nasal rather than the oral
route for respiration then do living humans. These specializations were very
possibly due to respiratory-related adaptations to their environment. A
by-product of this respiratory-driven anatomical configuration would be that
Neanderthals could not have produced the same array of sounds that living
humans can (16, 17, 26). They were not apish mutes; they were just not
identical to us.
Although the
above scenario has been accepted by many, it is unpalatable to some. This group,
often called ‘‘lumpers,’’ is primarily comprised of those who view
Neanderthals as falling within the range of variation represented by diverse
modern human populations (27). Given their predilection, it becomes a priori
impossible for them to view Neanderthals as ever being sufficiently
different so as to exhibit highly derived respiratory anatomy or specialized
respiratory or vocal behaviors. If they are us, then they cannot be
fundamentally different. Observations on the difference between Neanderthals
and extant populations are routinely dismissed as being within the range of ‘‘human’’
variation.
Given
the above, Schwartz and Tattersall’s nasal findings harbor important
implications. The three traits they describe clearly suggest a morphology that
is different from ours and appears designed to subserve specialized functions.
The large internal nasal margin they describe, their major observation, may
serve to expand the internal surface area, thus allowing for an increase in
ciliated mucosal covering. The placement of this margin, at the very entrance
of the cavity, also suggests a location ideally suited to be the initial
vehicle to confront
inspired air or
the last opportunity to interact with expired air.
In some ways,
this medial projection is reminiscent of Waldeyer’s Ring, the
mucosa-associated lymphoid tissues (lingual, palatine, nasopharyngeal tonsils)
that surround internal respiratory and digestive portals. Could Neanderthals
have had a similar ‘‘donut-shaped’’ filter system at the entrance to the
nasal cavity?
Both the internal nasal
margin and Schwartz and Tattersall’s second feature, a swelling of the lateral
nasal cavity, may be related to expansions of the paranasal sinus system.
Indeed, sinuses have long been known to be extensive in Neanderthals (28),
and amplifications of these would thus be logical. Al-though the exact
function(s) of mammalian paranasal sinuses remains unclear, and have indeed
become the focus of much recent study (29–31), it is likely that in
Neanderthals they played at least some part in an air-exchange process, perhaps
in warming and humidifying cold and dry air. The relationships/ functions of an
unossified roof over the lacrimal groove, Schwartz and Tattersall’s third
observation, is less clear. The absence of a rigid roof, however, would clearly
permit more expandability for components of the nasolacrimal duct system
(which, in humans, contains a venous plexus forming erectile tissues and can,
when engorged, obstruct the duct).
Absence of a bony roof
would also allow for a more direct communication of nasolacrimal duct contents
with the environment of the nasal cavity proper.
As an initial
foray into this region in Neanderthals, many questions have yet to be
addressed. An important issue, not covered by Schwartz and Tattersall, regards
the extent (or even presence) of these proposed apomorphies in Neanderthals
from different geographic areas. For example, because the Neanderthals focused
on by the authors are from western Europe, it will be important to determine the
extent of these characters in Neanderthals from other areas, such as eastern
Europe or the Levant. If regional variations are found, could there be any
relationship to climatic differences? Similarly, further work is needed to
determine...
whether, as the authors suggest, the presence of a ‘‘poorly developed’’
medial projection in the Steinheim cranium (a specimen
generally
thought not to be a Neanderthal) is evidence for an entire Neanderthal clade.
The
apomorphies described by Schwartz and Tattersall offer strong evidence that the
internal morphology of the nasal moiety of the Neanderthal upper respiratory
tract may be as distinctive as that previously reconstructed for the more
caudal laryngo-pharyngeal component. Taken together, these specialized
features of
different upper respiratory tract compartments allow greater insight into an
apparently more global pattern to Neanderthal upper respiratory
specializations. Although these specializations may not by themselves validate
Schwartz and
Tattersall’s preference for Neanderthal taxonomic distinctiveness, they
certainly lend strong support to those who see Neanderthals as considerably
different from living Homo sapiens. Indeed, further clues to
understanding just how different Neanderthals are from living humans may be
as plain as the
anatomy inside the noses on their faces.”
References:
1. Tattersall,
I. (1995) The Last Neanderthal: The Rise, Success, and
Mysterious
Extinction of Our Closest Human Relatives (Macmillan,
New York).
2. Stringer, C.
& Gamble, C. (1993) In Search of the Neanderthals:
Solving the
Puzzle of Human Origins. (Thames & Hudson, London).
3. Schwartz, J.
H. & Tattersall, I. (1996) Proc. Natl. Acad. Sci. USA
93, 10852–10854.
4. Schwartz, J.
H. (1993) What the Bones Tell Us (Henry Holt, New
York).
5. Tattersall,
I. (1986) J. Hum. Evol. 15, 165–175.
6. Thomson, A.
& Buxton, L. H. D. (1923) J. R. Anthropol. Inst. 53,
92–133.
7. Wolpoff, M.
H. (1968) Am. J. Phys. Anthropol. 29, 405–423.
8. Glanville,
E. V. (1969) Am J. Phys. Anthropol. 30, 29–38.
9. Franciscus,
R. G. & Trinkaus, E. (1988) Am. J. Phys. Anthropol.
75, 517–527.
10. Coon, C. S.
(1962) The Origin of Races (Knopf, New York).
11. Franciscus,
R. G. & Trinkaus, E. (1988) Am. J. Phys. Anthropol.
75, 208.
12. Trinkaus,
E. & Shipman, P. (1993) The Neandertals: Changing the
Image of
Mankind (Knopf, New York).
13. Laitman, J.
T., Heimbuch, R. C. & Crelin, E. S. (1978) Am. J.
Anat. 152, 467–483.
14. Reidenberg,
J. S. & Laitman, J. T. (1991) Anat. Rec. 230, 557–569.
15. Laitman, J.
T., Reidenberg, J. S. & Gannon, P. J. (1992) in
Language
Origin: A Multidisciplinary Approach, eds. Wind, J.,
Chiarelli, B.,
Bichakjian, B. & Nocentini, A. (Kluwer, Dordrecht,
The
Netherlands), pp. 385–397.
16. Laitman, J.
T., Heimbuch, R. C. & Crelin, E. S. (1979) Am. J.
Phys.
Anthropol. 51, 15–34.
17. Lieberman,
P. & Crelin, E. S. (1971) Ling. Inq. 2, 203–222.
18. Grosmangin,
C. (1979) Mem. Lab. Anat. Fac. Med. Paris 40,
1–241.
19. Budil, I.
(1994) Hum. Evol. 9, 35–52.
20. Laitman, J.
T. & Reidenberg, J. S. (1993) Dysphagia 8, 318–325.
21. Harrison,
D. F. N. (1995) The Anatomy and Physiology of the
Mammalian
Larynx (Cambridge Univ. Press, Cambridge, U.K.).
22. Laitman, J.
T. & Heimbuch, R. C. (1982) Am. J. Phys. Anthropol.
59, 323–344.
23. Negus, V.
E. (1949) The Comparative Anatomy and Physiology of
the Larynx (Grune &
Stratton, New York).
24. Laitman, J.
T., Reidenberg, J. S., Friedland, D. R., Reidenberg,
B. E. &
Gannon, P. J. (1993) Am. J. Phys. Anthropol. Suppl. 16,
129.
25. Crelin, E.
S. (1987) The Human Vocal Tract: Anatomy, Function,
Development and
Evolution (Vantage, New York).
26. Lieberman,
P., Laitman, J. T., Reidenberg, J. S. & Gannon, P. J.
(1992) J.
Hum. Evol. 23, 447–467.
27. Wolpoff, M.
H. (1996) Human Evolution (McGraw–Hill, New
York).
28. Tillier, A.
M. (1975) The`se de 3 e cycle (Univ. of Paris, Paris).
29. Scharf, K.,
Lawson, W., Shapiro, J. & Gannon, P. J. (1994)
Laryngoscope 105, 1–5.
30. Koppe, T.,
Rohrer-Ertl, O., Hahn, D., Reike, R. & Nagai, H.
(1996) Okajimas
Folia Anat. Jpn. 72, 297–306.
31. Marquez,
S., Gannon, P. J., Reidenberg, J. S. & Laitman, J. T.
(1996) Assoc.
Res. Otolaryngol. Abstr. 19, 158.
This original
comment can be found only in PDF for free at:
APPENDIX:
Some
mitochondrial DNA fragments from neandertals:
In the next address can be freely found this and any other
sequence from living beings:
(http://www.ncbi.nlm.nih.gov/Database/index.html)
AF011222
Krings,M., Stone,A., Schmitz,R.W., Krainitzki,H., Stoneking,M. and Paabo,S., Neandertal DNA sequences and the origin of modern humans, Cell 90 (1), 19-30 (1997)
Homo sapiens neanderthalensis
mitochondrial D-loop hypervariable region 1
1 gttctttcat gggggagcag atttgggtac cacccaagta ttgactcacc catcagcaac 61 cgctatgtat ctcgtacatt actgttagtt accatgaata ttgtacagta ccataattac 121 ttgactacct gcagtacata aaaacctaat ccacatcaaa cccccccccc catgcttaca 181 agcaagcaca gcaatcaacc ttcaactgtc atacatcaac tacaactcca aagacgccct 241 tacacccact aggatatcaa caaacctacc cacccttgac agtacatagc acataaagtc 301 atttaccgta catagcacat tacagtcaaa tcccttctcg cccccatgga tgacccccct361 cagatagggg tcccttgat
AF142095
Krings,M., Geisert,H., Schmitz,R.W., Krainitzki,H. and Paabo,S., DNA sequence of the mitochondrial hypervariable region II from the neandertal type specimen, Proc. Natl. Acad. Sci. U.S.A. 96 (10), 5581-5585 (1999).
Homo sapiens neanderthalensis mitochondrial control region, hypervariable region II. 1 ttttcgtctg gggggtgtgc acgcgatagc attgcgagac gctggagccg gagcacccta 61 tgtcgcagta tctgtctttg attcctgccc cattccatta tttatcgcac ctacgttcaa 121 tattacaggc gagcatactt actaaagtgt gttaattaat taatgcttgt aggacataat 181 aataacgact aaatgtctgc acagctgctt tccacacaga catcataaca aaaaatttcc 241 accaaacccc ctttcctccc ccgcttctgg ccacagcact taaacacatc tctgccaaac301 cccaaaaaca aagaacccta acaccagcct aaccagactt caaat
AF254446
Ovchinnikov,I.V., Gotherstrom,A., Romanova,G.P., Kharitonov,V.M., Liden,K. and Goodwin,W., Molecular analysis of Neanderthal DNA from the northern Caucasus, Nature 404 (6777), 490-493 (2000).
Homo sapiens neanderthalensis mitochondrial D-loop, hypervariable region I. 1 ccaagtattg actcacccat caacaaccgc catgtatttc gtacattact gccagccacc 61 atgaatattg tacagtacca taattacttg actacctgta atacataaaa acctaatcca 121 catcaacccc ccccccccat gcttacaagc aagcacagca atcaaccttc aactgtcata 181 catcaactac aactccaaag acacccttac acccactagg atatcaacaa acctacccac 241 ccttgacagt acatagcaca taaagtcatt taccgtacat agcacattat agtcaaatcc301 cttctcgccc ccatggatga cccccctcag ataggggtcc cttga
AF282971
Krings,M., Capelli,C., Tschentscher,F., Geisert,H., Meyer,S., von Haeseler,A., Grossschmidt,K., Possnert,G., Paunovic,M. and Paabo,S., A view of neandertal genetic diversity, Nat. Genet. 26 (2), 144-146 (2000).
Homo sapiens neanderthalensis mitochondrial hypervariable region I sequence. 1 gttctttcat gggggagcag atttgggtac cacccaagta ttgactcacc catcagcaac 61 cgctatgtat ttcgtacatt actgccagcc accatgaata ttgtacagta ccataattac 121 ttgactacct gcagtacata aaaacctaat ccacatcaac cccccccccc catgcttaca 181 agcaagcaca gcaatcaacc ttcaactgtc atacatcaac tacaactcca aagacgccct 241 tacacccact aggatatcaa caaacctacc cacccttgac agtacatagc acataaagtc 301 atttaccgta catagcacat tacagtcaaa tcccttctcg cccccatgga tgacccc AF282972
Krings,M., Capelli,C., Tschentscher,F., Geisert,H., Meyer,S., von Haeseler,A., Grossschmidt,K., Possnert,G., Paunovic,M. and Paabo,S., A view of neandertal genetic diversity, Nat. Genet. 26 (2), 144-146 (2000).
Homo sapiens neanderthalensis mitochondrial hypervariable region II sequence. 1 ttttcgtctg gggggtgtgc acgcgatagc attgcgagac gctggagccg gagcacccta 61 tgtcgcagta tctgtctttg attcctgccc cattccatta tttatcgcac ctacgttcaa 121 tattacaggc gagcatactt actgaagtgt gttaattaat taatgcttgt aggacataat 181 aataacgact aaatgtctgc acagctgctt tccacacaga catcataaca aaaaatttcc
241 accaaacctc cccctccccc gcttctggcc acagcactta aatacatc
AY149291
Schmitz,R.W., Serre,D., Bonani,G., Feine,S., Hillgruber,F., Krainitzki,H., Paabo,S. and Smith,F.H., The Neandertal type site revisited: interdisciplinary investigations of skeletal remains from the Neander Valley, Germany, Proc. Natl. Acad. Sci. U.S.A. 99 (20), 13342-13347 (2002).
Homo sapiens neanderthalsensis mitochondrial D-loop hypervariable region I, partial sequence. 1 gttctttcat gggggagcag atttgggtac cacccaagta ttgactcacc catcagcaac 61 cgctatgtat ttcgtacatt actgccagcc accatgaata ttgtacagta ccataattac 121 ttgactacct gcagtacata aaaacctaat ccacatcaac cccccccccc catgcttaca 181 agcaagcaca gcaatcaacc ttcaactgtc atacatcaac tacaactcca aagacaccct
241 tacacccact aggatatcaa caaacctacc cacccttgac agtacatagc acataaagtc 301 atttaccgta catagcacat tacagtcaaa tcccttctcg cccccatgga tgacccc
Comparing Neanderthal sequences with human's (with "BLAST"), and also human - human comparisons:
http://www.oocities.org/fdocch/compare.htm
Go to the first part of this review:
http://www.oocities.org/fdocch/new.htm
More Neanderthal mitochondrial DNA Research:
http://www.oocities.org/fdocch/neil.htm
Clear proofs of Neandertal Cannibalism: Cave finds revive Neandertal cannibalism (Evidence found at cave site in southeastern France):
Science News, Oct 2, 1999, B. Bower
Neanderthal Eats Neanderthal:
Neandertals' diet put meat in their bones:
Science News June 17, 2000, B. Bower
Neanderthals Did Their Own Killing:
By Trina Wood, Discovery.com News, June 13, 2000
White, Tim D. (August, 2001). “Once Were Cannibals"Scientific American 58-65.
An excerpt in Word Document of this thoughtful article
Some other references of the neandertal cannibalism:
Defleur, A., T. White, et al. (1999). "Neanderthal cannibalism
at Moula-Guercy, Ardeche, France." Science 286(5437): 128-31.
Richards, M. P., P. B. Pettitt, et al. (2000). “Neanderthal diet
at Vindija and Neanderthal predation: the evidence from stable isotopes.” Proc
Natl Acad Sci U S A 97(13): 7663-6.
White, T. D. (1992). “Prehistoric Cannibalism at Mancos 5MTUMR-2346."Princeton University Press
Marlar, R. A., et al (September 7, 2000). “Biochemical Evidence of Cannibalism at a Prehistoric Puebloan Site in Southwestern Colorado."Nature (407):74-78.
http://www.webster.sk.ca/GREENWICH/chewchip.htm CHEWED OR CHIPPED? Who Made the Neanderthal Flute? HUMANS OR CARNIVORES?
By Bob Fink
Updated Nov, 2ooo
New attempts made by a Scientific foolishness: the "reproduction" of two individuals of the same sex: "Theoretically, these new techniques could eventually also allow
same-sex couples to reproduce", according to the article of Peter O'Connor
... this example and any other homosexually oriented program, are wrong, according to God's Word:
New Experimental Fertility Technique
Tasters of the Word (YouTube), videos recientes: "Astronomía y Nacimiento de Jesucristo: Once de Septiembre Año Tres A.C.", "Estudio sobre Sanidades" (en 20 episodios), "Jesus Christ, Son or God?" and "We've the Power to Heal":
Tasters of the Word (the blog, with: "Astronomy and the Birth of Jesus Christ"):